- The paper demonstrates that activity-dependent plasticity significantly boosts performance in competent morphogenetic networks, yielding over +60 reward gains on CartPole.
- The paper shows that anti-Hebbian plasticity consistently outperforms Hebbian updates, with statistically significant improvements (Cohen’s d=0.53–0.64) across tasks.
- The paper reveals that co-evolving plasticity parameters with developmental ones leads to task-specific, per-network tuning that effectively adapts to dynamic environments.
Activity-Dependent Plasticity in Morphogenetically-Grown Recurrent Networks: A Technical Analysis
Introduction and Motivation
This paper explores the intersection of developmental neural network encodings and activity-dependent synaptic plasticity, focusing on task-driven adaptation in networks produced by morphogenetic self-organization. Specifically, the work leverages MorphoNAS—a system that generates network architectures via Turing-style reaction-diffusion morphogen gradients—to investigate how simple synaptic plasticity rules (Hebbian and anti-Hebbian) interact with diverse, developmentally grown recurrent networks. The central questions are: (1) Under what circumstances does activity-dependent plasticity enhance the functionality of such evolved networks? (2) Can evolutionary processes optimize plasticity parameters in synergy with developmental encoding, and is this interaction fundamentally different from that in random networks?
Morphogenetic Development and Evaluation Protocol
MorphoNAS leverages a compact, 54-parameter genome specifying morphogen dynamics (secretion, diffusion, inhibition), cell-fate decisions, and chemotactic axon growth. The deterministic developmental procedure begins with a single cell on a 2D grid; reaction-diffusion gradients induce local rules for division, differentiation, and axonal extension, giving rise to topologically and functionally diverse recurrent neural networks (RNNs). This protocol enables the rapid sampling of tens of thousands of unique architectures, far exceeding the expressiveness of direct neural encoding.

Figure 1: Developmental dynamics of a MorphoNAS network on a 10×10 grid, where morphogen concentrations drive neuronal proliferation and connectivity.
Performance was assessed on CartPole-v1 and Acrobot-v1, classic reinforcement learning benchmarks. Topologies were stratified by baseline CartPole reward (without plasticity): the vast majority produced non-viable ("Weak") controllers, highlighting developmental structure as the dominant determinant of function.
Systematic Characterisation of Synaptic Plasticity
The paper introduces a rigorous, population-scale analysis of the effects of Hebbian and anti-Hebbian plasticity, parameterized by a learning rate η and decay λ, according to:
Δwij=η⋅xixj−λwij
Grid sweeps of (η,λ) over 50,000 networks yielded more than five million configuration evaluations.
Key empirical findings:
- Anti-Hebbian plasticity is consistently superior for competent developmental networks. Fixed-parameter oracle tuning shows that up to 93% of high-performing networks benefit, with Cohen’s d up to 0.64 in favor of anti-Hebbian synaptic updates.
- Plasticity’s utility is strictly topology-limited: Plasticity cannot salvage nonfunctional (weak) networks, implying functional competence must be bootstrapped by development.
- Parameter heterogeneity is extreme: No universal (η,λ) maximizes performance across topologies. Regret, defined as the fraction of oracle improvement lost by using the globally best setting, ranges from 52% to 100%, particularly acute for developmentally generated networks.
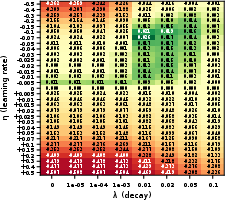
Figure 2: High-mid CartPole networks achieve maximal gains for anti-Hebbian (η<0) with moderate decay (λ=0.01); Hebbian updates are deleterious.
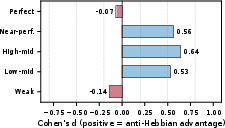
Figure 3: Effect sizes (Cohen’s d) confirm anti-Hebbian dominance for all competent strata (d = 0.53–0.64; η0).
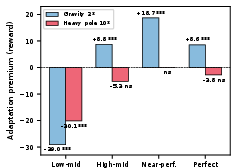
Adaptation Under Environmental Nonstationarity
Plasticity provides qualitatively distinct benefits when networks must adapt to sudden environmental changes. Evaluation under physics perturbations (pole mass ×10, gravity ×2) shows that:
A targeted OFFη4ON protocol, activating plasticity only after the environment shift, isolates adaptation: most of the plasticity benefit is retained, scaling with the adaptation window.
Generalisation to Acrobot and Topology-Plasticity Interactions
Acrobot-v1 experiments, with an expanded developmental grid, reproduce the key pattern: over 93% of networks benefit from per-topology oracle tuning, but again, fixed parameter settings perform suboptimally. There is a marked reduction in optimal η5 magnitude (requiring η6–η7 lower rates versus CartPole), and the advantage of anti-Hebbian over Hebbian is less uniform (Cohen’s η8–η9). Regret remains at 100% under any fixed setting.
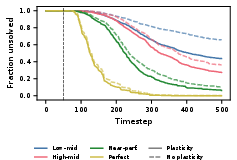
Figure 5: Fraction of unsolved Acrobot episodes is reduced by per-network oracle plasticity, especially pronounced under environmental change at step 50.
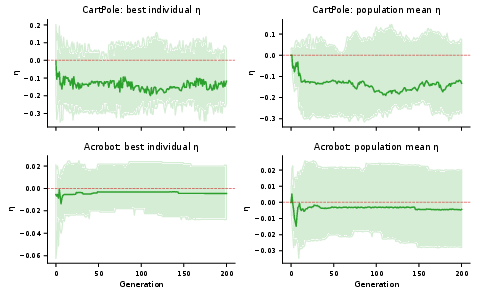
Co-Evolution of Developmental and Plasticity Parameters
A genetic algorithm was used to evolve both morphogenetic parameters and plasticity rule parameters. The principal observations:
Disentangling Developmental and Generic Network Effects
Random-RNN controls, matched for size and density to MorphoNAS outputs, establish that:
- Morphogenetic development imparts an λ4 higher probability of sampling competent controllers than random wiring.
- Anti-Hebbian dominance is generic to small RNNs controlling CartPole but the dependence on topology (i.e., per-network regret) is far greater for morphogenetic networks. Plasticity’s interaction with architecture is more variable in developmentally constructed networks, making co-optimisation with plasticity much more significant.
Implications and Future Directions
Theoretical Implications: The work empirically substantiates the conjecture that genomic bottleneck mechanisms, as present in biological systems, play a crucial role in constraining the functional space toward architectures that are both compact and adaptive. Morphogenetic development robustly bootstraps viable computational structure, while simple anti-Hebbian plasticity rules optimize local adaptation. However, the necessity of individualized plasticity parameterization for each developmental phenotype suggests a limit to general-purpose plasticity rule design in highly variable topological regimes.
Practical Implications: For practitioners seeking to deploy self-organizing neural systems in dynamic environments, the optimal protocol is: (1) use anti-Hebbian plasticity with moderate decay as a safe default, (2) perform per-network parameter optimization whenever possible, and (3) rely on environment-triggered plasticity for non-stationary settings.
Limitations and Prospects: The findings are robust on small-scale discrete-action control tasks; extension to continuous-action, high-dimensional, or multi-agent domains is an open research direction. The observed counterproductivity of co-evolving plasticity under certain conditions suggests the need for richer plasticity mechanisms and more complex selection environments.
Conclusion
This paper precisely characterizes how synaptic plasticity rules interact with morphogenetically generated network architectures, demonstrating that anti-Hebbian plasticity typically dominates in functional, small recurrent controllers. The role of plasticity transitions from static fine-tuning to essential adaptation under non-stationarity, but topological diversity from development provokes extreme heterogeneity in plasticity parameter optima. Evolutionary search, when co-optimizing architecture and plasticity, recapitulates these findings, but the developmental process, not plasticity alone, is the dominant source of functional competence. The results provide a comprehensive empirical and methodological foundation for future research at the intersection of self-organizing neural development and adaptive computation in artificial intelligence systems.
Reference:
"Activity-Dependent Plasticity in Morphogenetically-Grown Recurrent Networks" (2604.03386)